
ES細胞とiPS細胞:幹細胞のあれこれ
はじめに
大学での授業アンケートで,「授業中に話題に取り上げなかったことで,もっと知りたかったことはありますか?」と聞くと,必ず「STAP細胞はありますか?」と質問されます.手短に答えると,STAP(stimulus triggered acquisition of pluripotency)細胞と命名された全能性細胞は存在しません.しかし,STAP細胞よりも前に,比較的簡単に多分化能を獲得できる細胞が大人の骨髄などに含まれていることが知られていました.この細胞集団を体から取り出して培養すると,間葉系幹細胞(MS cells: mesenchymal stem cells)(文献1, 2)とミューズ細胞(Muse cells: multi-lineage differentiating stress enduring cells)(文献3, 4)と呼ばれる2種類の幹細胞を得ることができます.私が担当する生物学の新知識は,表1に示す14項目からなっています.この幹細胞シリーズを通して,「夢の幹細胞」とは存在するのか?そもそも幹細胞は難病患者を救えるのか?なぜiPS細胞が重要なのか?無制限にiPS細胞を利用していいのか?といった疑問を,読者のみなさんがご自身で判断できるよう,基礎知識を共有していきたいと思います.

1. 幹細胞(stem cells)ってなに?
1–1. 幹細胞とは
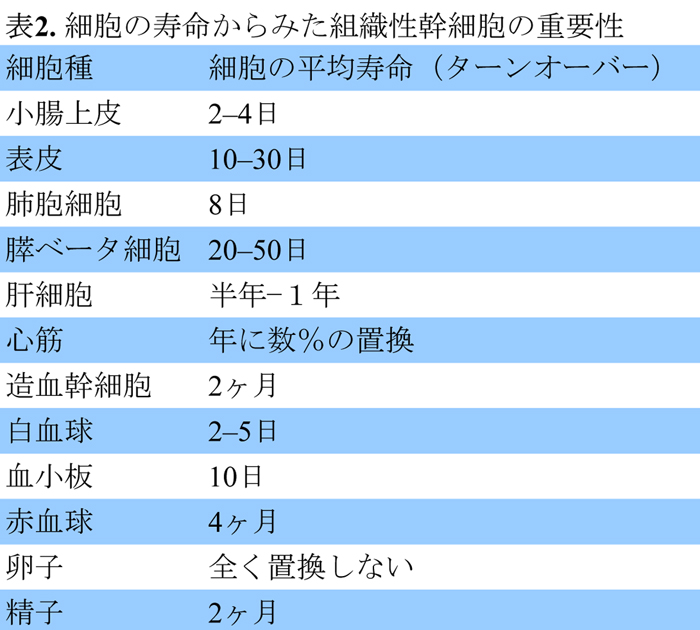
私たちの体はたくさんの種類の細胞(体細胞)によって形作られ,それらがもつ固有の機能が寄り集まって恒常性を維持しています.体細胞には寿命があり,役割を終えた細胞は新たな細胞に置き換えられます.そのスピードは組織によって異なっていて,ものすごく置換の早い小腸上皮細胞の寿命は,たったの1日か2日しかありません.大きな臓器でサイズの変化がない肺,肝臓や膵臓などでも,細胞は頻繁に交換されています(表2)(文献5).特に,体外から取り入れた化合物を解毒する機能をもつ肝臓では,肝細胞が障害を受けるやいなや細胞数が激減しますが,通常3日程度で新しい肝細胞で修復されます.一方,心臓を動かしている心筋や卵巣に含まれる生殖細胞の卵子は,生まれた後に増えません.このことは,あらゆる組織には,固有の特性をもつ細胞供給源があることを示しています.組織の中で生命活動の維持に必要な体細胞を供給している細胞を,体性幹細胞または組織性幹細胞といいます.
幹細胞とは,必要に応じて分裂を繰り返すことができる自己複製能(self-renewal potency)と機能的に分化した細胞を供給できる分化能(differentiation potency)を合わせ持つ細胞です.例えば,骨髄には造血幹細胞と呼ばれる体性幹細胞が存在し,自己増殖しながらも白血球や赤血球,血小板などさまざまな血球系細胞を絶え間なく供給しています.造血幹細胞は,骨髄移植によって他人の体に注入されても,状況が整うと増えて血球系細胞を供給します (図1).

図1. 体細胞を絶えず供給する体性幹細胞
幹細胞は,自己複製能と分化能を持ちます.
幹細胞は,自己複製能と分化能を持ちます.
再生医療とは,細胞を移植したり,体に残っている体性幹細胞を活性化させたりすることで,直接的・間接的に損なわれた組織の機能を回復させる医療を指します.つまり,本来,幹細胞を用いた細胞移植を前提としていません.他人の血液,上皮組織,目の角膜など,細胞移植治療は既に数多く実績を上げてきています.最近メディアで取り上げられている「再生医療」というキーワードは,特に,初期胚細胞に近い未熟な幹細胞から作り出した組織細胞を用いた細胞移植治療を指す傾向にあります.
1–2. 多能性幹細胞とは
未熟な発生段階に留まっている幹細胞の性質を「未分化性」といいます.未分化な幹細胞は,その分化能に応じて幾つかのクラスに分けられています.1個の細胞だけで個体を作りだせる幹細胞の能力は全能性(totipotency)と呼ばれ,高等動物では受精卵だけが持っています.一方,体を構成する全ての組織細胞や生殖細胞に分化できる未分化な幹細胞を,多能性幹細胞(pluripotent stem cells)と呼びます.体性幹細胞の多くは生殖細胞以外の組織細胞に分化できる限定的な多能性をもち,multipotent stem cellsに分類されています.pluripotent とmultipotentをうまく区別できる日本語がないため,前者を万能性,後者を多能性と呼ぶこともあります.iPS細胞(induced pluripotent stem cells)は,人工的に体細胞から誘導(induced)して作製した多能性幹細胞という意味を持ちます(文献6)(図2).

図2. ES細胞とiPS細胞
どちらも無限に分裂し,さまざまな組織細胞に分化できます.特に,ヒトiPS細胞は,体細胞ドナーにとって免疫拒絶のない移植細胞の供給源になると期待されています.
どちらも無限に分裂し,さまざまな組織細胞に分化できます.特に,ヒトiPS細胞は,体細胞ドナーにとって免疫拒絶のない移植細胞の供給源になると期待されています.
受精卵が発生を進めながら子宮に着床する頃になると,初期胚の中に体や生殖細胞を作る元になる多能性幹細胞が他の細胞から分かれ塊を形成します.この内部細胞塊細胞を培養皿に取り出して培養すると,長期間増殖しても多分化能を維持し続けられる幹細胞が現れます.これがES細胞(embryonic stem cells)です(文献7)(図2).マウスES細胞は分化能が高く,同等の発生段階にある初期胚に混ぜて擬妊娠マウスの子宮に戻すと,キメラマウスが生まれます.とりわけ,2細胞期胚の割球を融合して作った胎盤にしか分化できない4倍体胚にES細胞を混ぜて擬妊娠マウスの子宮に戻すと,100% ES細胞からできたマウスが誕生します.4倍体胚盤胞補完法と呼ばれるこの優れた胚操作技術を使って,マウスiPS細胞に完全な多能性があることが証明されています(文献8).
1–3. リプログラミング因子とは
ES細胞やiPS細胞は,その多分化能から,さまざまな組織細胞の大量供給源になると期待されています(文献9, 10).しかし,その実現には,ES細胞やiPS細胞の多分化能を失わないように,細心の注意を払って未分化性を維持しながら培養することが肝心です.では,ES細胞はどのように未分化性を維持しているのでしょうか?この未分化性維持に必要な仕組みを細胞の中で動かしているメインスイッチが,山中4因子の中の3因子Oct4,Sox2,Klf4なのです.
細胞の中には,さまざまなタンパク質が詰め込まれています.ヒト子宮頸がん細胞株HeLaの場合,1細胞当たり20億個のタンパク質を含んでいます(文献5).小さな大腸菌E. coliは,ヒトより少ない230万個のタンパク質を含みますが,体積当たりで比較すると,驚くことにヒトの2倍以上のタンパク質を含んでいます.量だけでなくタンパク質の種類が重要で,細胞種によって大きく異なる組成のタンパク質は,相互に作用し合って固有の細胞機能を制御しています.iPS細胞研究における最大の発見は,山中4因子と呼ばれる4つの遺伝子Oct4, Sox2, Klf4, cMycだけで,あらゆる体細胞型の分子ネットワークを抑制し,新たに多能性幹細胞型の分子ネットワークを再構築できることを見いだしたことにあります.メインスイッチが入って幹細胞型の遺伝子発現制御機構が働き出すと,外来性の山中4因子はもはや不要で,体細胞だったときに眠っていた遺伝子群が活発に未分化性を維持するようになります.このように発生を巻き戻して未分化に戻す因子を,リプログラミング因子または初期化因子と呼びます(図2).
細胞の中には,さまざまなタンパク質が詰め込まれています.ヒト子宮頸がん細胞株HeLaの場合,1細胞当たり20億個のタンパク質を含んでいます(文献5).小さな大腸菌E. coliは,ヒトより少ない230万個のタンパク質を含みますが,体積当たりで比較すると,驚くことにヒトの2倍以上のタンパク質を含んでいます.量だけでなくタンパク質の種類が重要で,細胞種によって大きく異なる組成のタンパク質は,相互に作用し合って固有の細胞機能を制御しています.iPS細胞研究における最大の発見は,山中4因子と呼ばれる4つの遺伝子Oct4, Sox2, Klf4, cMycだけで,あらゆる体細胞型の分子ネットワークを抑制し,新たに多能性幹細胞型の分子ネットワークを再構築できることを見いだしたことにあります.メインスイッチが入って幹細胞型の遺伝子発現制御機構が働き出すと,外来性の山中4因子はもはや不要で,体細胞だったときに眠っていた遺伝子群が活発に未分化性を維持するようになります.このように発生を巻き戻して未分化に戻す因子を,リプログラミング因子または初期化因子と呼びます(図2).
1–4. マウスES細胞の未分化性維持に必要なLIFとは
未分化状態に戻った多能性幹細胞は,分化関連遺伝子を抑制状態に,未分化性維持に関連する遺伝子を積極的かつ恒常的に活性化する仕組みを必要とします.そのため,山中4因子を発現する遺伝子を導入してiPS細胞を作製する時には,ES細胞を培養する条件で体細胞を培養します.マウスとヒトは,ES細胞やiPS細胞の性質が大きく異なり,その未分化性維持機構も違います(図3).本項では,マウスES細胞の未分化性維持機構を中心に記載します.

図3. マウスES細胞とヒトiPS細胞の違い
フィーダー細胞の上で培養したマウスES細胞とヒトiPS細胞は,細胞の形もコロニーの形態も違います.
フィーダー細胞の上で培養したマウスES細胞とヒトiPS細胞は,細胞の形もコロニーの形態も違います.
マウスES細胞の培養液には,白血病抑制因子(LIF:leukaemia inhibitory factor)を加えます.LIFは細胞外から細胞核内部まで「細胞よ,未分化になれ!」とシグナルを伝える魔法の粉(リガンド)として働き,2種類のタンパク質LIFRとgp130の組み合わせからなる受容体に結合し,少なくとも以下に上げる4種類のシグナル伝達経路を活性化します: ①JAK1/STAT3,②Ras/ERK1/2経路,③PI3K/Akt経路,④SFK経路.これらのシグナルは核内に伝えられ,ターゲット遺伝子の発現を制御します.中でも,①JAK1/STAT3シグナルは,Klf4発現を活性化し,Klf4はSox2発現の誘導とOct4発現の維持に働きます.②PI3K/Aktシグナルは,Tbx3の活性化を介してNanog発現を誘導します(図4).Nanogは,多能性幹細胞の維持にとても重要な転写因子です.私たちは,Nanogは胚発生過程で極めて未熟な幹細胞で特異的に発現し,発現量依存的に未分化状態の維持に働いていることを見いだしました(文献11).また,Oct4がSox2と協調してNanogを発現させることも明らかにています(文献12).そのNanogは,再びOct4発現を活性化することで,幹細胞の未分化性をより一層安定化します.このように,LIFは,ES細胞の未分化性の維持,幹細胞の生存,分化能の維持に必要なさまざまな遺伝子を制御するマスター因子Oct4/Sox2/NanogをKlf4の活性化を介して働かせることで,ES細胞特性を無限に維持することができるのです(文献13)(図5上).
山中4因子の残る1つcMycは,Maxとタンパク質複合体を形成し,ES細胞を分化させてしまうMAPKシグナルを抑制することで間接的に未分化性維持に働きます(文献14)(図5下).癌遺伝子として知られるcMycは,Klf4/Oct4/Sox2/Nanogの作用を高める働きをもつもののリプログラミング自体に必須ではありません.そこで,移植治療用iPS細胞の作製には,cMycを用いない方法が推奨されています.
-

図4. マウスES細胞の未分化性を維持するしくみと山中4因子との関係
マウスES細胞培養液にLIFを加えると山中4因子が動きだし,未分化性を維持します. -

図5. リプログラミング因子の働き
癌遺伝子cMycは,リプログラミングよりも分化抑制にはたらいています.
2. 体細胞から幹細胞へ発生を巻き戻す
iPS細胞は,ES細胞の多能性維持に働いている遺伝子群,通称ECAT (ES cell associated transcripts)を体細胞で発現させる実験に始まっています.なぜECATを用いたかというと,iPS細胞樹立以前から,未受精卵やES細胞には体細胞をリプログラミングできる初期化因子が含まれていることが知られていたからです.
1996年に世界初の体細胞核移植クローン動物である羊のドリーが誕生し,卵の細胞質に体細胞核をリプログラミングする因子が含まれていることが示されました(文献15).同じ頃,私たちは,ES細胞と等しい多分化能をもつ胎児生殖細胞由来の幹細胞であるEG細胞(embryonic germ cells)を体細胞と細胞融合すると,体細胞核のDNAのメチル化修飾がほとんど消えてしまうことを見いだしました(文献16).次に,マウスES細胞と体細胞を細胞融合して,体細胞核のDNAメチル化修飾が消えるだけでなく,ヒストンタンパク質にDNAが巻き付く強さを制御しているタンパク質修飾が固い体細胞型から緩いES細胞型に書き換えられることを見いだしました(図6)(文献17, 18).DNAやヒストンタンパク質に付けられている書き換え可能な遺伝子発現のプログラムをエピジェネティクスといいます。私たちは,このように細胞融合実験をもちいて,世界で始めて遺伝子発現のプログラムが書き換えられるエピジェネティクスの再プログラム化(リプログラミング)現象を目で捉えられる形で証明しました.同時に,ES細胞の中ではリプログラミング因子が常に発現していることを示したのです(図6).
1996年に世界初の体細胞核移植クローン動物である羊のドリーが誕生し,卵の細胞質に体細胞核をリプログラミングする因子が含まれていることが示されました(文献15).同じ頃,私たちは,ES細胞と等しい多分化能をもつ胎児生殖細胞由来の幹細胞であるEG細胞(embryonic germ cells)を体細胞と細胞融合すると,体細胞核のDNAのメチル化修飾がほとんど消えてしまうことを見いだしました(文献16).次に,マウスES細胞と体細胞を細胞融合して,体細胞核のDNAメチル化修飾が消えるだけでなく,ヒストンタンパク質にDNAが巻き付く強さを制御しているタンパク質修飾が固い体細胞型から緩いES細胞型に書き換えられることを見いだしました(図6)(文献17, 18).DNAやヒストンタンパク質に付けられている書き換え可能な遺伝子発現のプログラムをエピジェネティクスといいます。私たちは,このように細胞融合実験をもちいて,世界で始めて遺伝子発現のプログラムが書き換えられるエピジェネティクスの再プログラム化(リプログラミング)現象を目で捉えられる形で証明しました.同時に,ES細胞の中ではリプログラミング因子が常に発現していることを示したのです(図6).

図6. 細胞融合が教えてくれたES細胞のリプログラミング活性の存在
ES細胞と体細胞の融合細胞は,多能性をもつES細胞のような細胞です。
ES細胞と体細胞の融合細胞は,多能性をもつES細胞のような細胞です。
融合細胞は,ES細胞と体細胞からの染色体をもつ4倍体(4n)細胞ですが,ES細胞と同じようにさまざまな組織細胞を作り出すことができる分化多能性を持っています.そこで,免疫拒絶に関わるHLA遺伝子群を除去したES細胞を予め準備しておくと,融合細胞は体細胞ドナーのHLAだけを発現する半自己細胞を無限に提供することが原理的に可能になるのです(図6).
2–1. 細胞融合が解き明かしたES細胞のリプログラミング活性
私たちは,この2001年より体細胞を初期化する因子を探索する研究をスタートしました.EG細胞はES細胞よりも体細胞核のエピジェネティクスを消す能力が高かったことから,2種類の細胞の遺伝子発現を比較するcDNAサブトラクション法を行い,EG細胞でES細胞よりも2倍以上高く発現し,体細胞で発現していない遺伝子を探索しました.幹細胞特異的に発現している遺伝子にstem cellsを短縮したStmという名前を付けて解析を進めていました.この遺伝子の機能を解析していた頃,Nanog/ECAT4が発表され,Stmと同じ遺伝子であることが判明しました.このため,1-4項に前述したNanog遺伝子の特性は,Stm遺伝子の解析結果でした.私たちがNanog遺伝子クローニングに行き着いたように,細胞融合実験は,誰もが手にすることができるES細胞の中にリプログラミング因子があることを示したことで,世界中にリプログラミング因子探索の手がかりを提供することに繋がりました.
2–2. リプログラミングの解析技術開発の試み
私たちは,リプログラミング現象を捉えるためのさまざまな工夫をしてきました.今となっては,この技術開発が研究の中で最も楽しかった部分で,その後も沢山の研究に活用されています.まず,体細胞が未分化に戻るとES細胞で働いている遺伝子が働きだすことから,Oct4遺伝子の発現制御領域の下流に緑に光る蛍光タンパク質をコードするEGFP遺伝子を連結したOct4-EGFPレポーターをもつマウス体細胞を利用することを思い付きました.このOct4-EGFPトランスジェニックマウスは,共同研究をしていた理化学研究所の阿部訓也先生が開発したものです.Oct4-EGFPは体細胞では抑制されていているため,光っていない体細胞とES細胞を融合すると,48時間以内に光りだしました.同様のリプログラミング可視化システムは,さまざまなリプログラミング因子探索の効率化に貢献しました.
リンパ球には,さまざまな種類の抗体や細胞表面抗原を1対の遺伝子から作りだすため,遺伝子のDNAを少しずつ切り出してつなげるV(D)J組換えを起こしていることが知られています.ES細胞には,このDNA組換えの痕跡はありません.この情報は,共同研究をした徳島大学の高浜洋介先生に教えて頂きました.Oct4-EGFPマウスのリンパ球をES細胞と細胞融合し緑に光っている多能性の融合細胞のDNAを調べると,V(D)J組換えの痕跡が見つかるのです.リプログラミングされた細胞が本当に分化していた細胞だったのかを調べる方法として,今でも活用されています.この方法が,Oct4-EGFPと共にSTAP細胞の証明に活用されたことは,とても,とても複雑な気持ちです.
融合細胞の中では,体細胞とES細胞に由来するDNAやRNAが混在した状態になります.ES細胞と遺伝的に離れた種や系統の体細胞を用いると,DNA配列の違いから容易に2者を識別同定できるようになります.当時,ES細胞とは異なる亜種のJF1マウスが国立遺伝学研究所で繁殖され始めていました.私たちは,JF1マウスの体細胞を細胞融合に用いて,融合細胞のDNAや転写されているRNAの配列を解析しました.これにより,ES細胞のリプログラミング活性は十分に高く,リプログラミングされた体細胞核はES細胞の核と同等に変化し,遺伝子が同等に使われていることをようやく示すことができました(文献18, 19).
リンパ球には,さまざまな種類の抗体や細胞表面抗原を1対の遺伝子から作りだすため,遺伝子のDNAを少しずつ切り出してつなげるV(D)J組換えを起こしていることが知られています.ES細胞には,このDNA組換えの痕跡はありません.この情報は,共同研究をした徳島大学の高浜洋介先生に教えて頂きました.Oct4-EGFPマウスのリンパ球をES細胞と細胞融合し緑に光っている多能性の融合細胞のDNAを調べると,V(D)J組換えの痕跡が見つかるのです.リプログラミングされた細胞が本当に分化していた細胞だったのかを調べる方法として,今でも活用されています.この方法が,Oct4-EGFPと共にSTAP細胞の証明に活用されたことは,とても,とても複雑な気持ちです.
融合細胞の中では,体細胞とES細胞に由来するDNAやRNAが混在した状態になります.ES細胞と遺伝的に離れた種や系統の体細胞を用いると,DNA配列の違いから容易に2者を識別同定できるようになります.当時,ES細胞とは異なる亜種のJF1マウスが国立遺伝学研究所で繁殖され始めていました.私たちは,JF1マウスの体細胞を細胞融合に用いて,融合細胞のDNAや転写されているRNAの配列を解析しました.これにより,ES細胞のリプログラミング活性は十分に高く,リプログラミングされた体細胞核はES細胞の核と同等に変化し,遺伝子が同等に使われていることをようやく示すことができました(文献18, 19).
おわりに
私たちは,クローン羊のドリー誕生に始まるリプログラミング研究分野のスタート地点に遭遇し,リプログラミング現象を捉えて証明する方法の開発をしながら手探り状態で進んできました.当時、DNAやヒストン修飾の役割,それらを制御する酵素や関連因子が次々と明らかになってきていたことが解析の幅を広げました.何と言っても私たちの最大のリプログラミング解析ツールは「細胞融合法」でした.ヒントになった細胞融合実験は,テラトーマと呼ばれる良性の胚性ガン細胞(EC細胞:embryonal carcinoma cells)のPSAやOTF9とマウス体細胞の融合実験です.北海道大学大学院博士後期課程でご指導頂いた高木信夫先生は,とてもとても若かった頃にEC細胞と雌体細胞を細胞融合すると不活性X染色体が再活性化するという発見をしました(文献20).雌マウスの細胞にはX染色体が2本(XX)含まれていて,未分化細胞では2本とも活性化しているのに対し,胚が子宮に着床する頃になるとXY雄細胞との遺伝子量を補正する機構が働き出し,1本のX染色体が不活性化します.大学院生の私の頭にひらめいたのは,体細胞の不活性X染色体がEC細胞融合によって再活性化するのは,EC細胞には体細胞核を未分化状態に引き戻す活性があるからではないかということでした.X染色体の活性状態や別の機会にご紹介するインプリント遺伝子のDNAメチル化状態は,発生段階によって変化することから,今もOct4-EGFPと同様にリプログラミングを示すマーカーとして使われています.高木先生のお陰でX染色体の再活性化とEC細胞融合に出会えたのが,私のリプログラミング研究のはじまりのはじまりです.
次回の予告
マウスに続きヒトiPS細胞が樹立され,臨床応用も開始されています.ヒトiPS細胞は,マウスと同じ山中4因子または癌関連遺伝子であるcMycを除いた4つの因子OCT4, SOX2, NANOG, LIN28などを用いてヒトの皮膚細胞などから2007年に樹立されています(文献9, 10).興味深いことに,マウスとヒトでは,ES細胞の未分化性の維持機構が異なり,ヒトES細胞はマウスよりやや発生の進んだ状態にあります(図3).そのため,ヒトES細胞やiPS細胞の培養には,LIFではなくbFGF(別名FGF2)を培養液に加える必要があります(文献21).サルやヒトES細胞の未分化性維持機構が予め明らかになっていなければ,ヒトiPS細胞の樹立は困難だったと言えます.
現在,私たちは,ヒトiPS細胞から創薬開発に有用な良質のヒト肝細胞を代替する細胞を沢山作り出すことを目指しています.マウスやヒトのES細胞で既に膨大な分化研究がなされているものの、その品質は生体肝細胞からは遥かに遠いものに留まっています.溢れるほどの情報を活用しながらも,組織細胞が機能的に成熟するとはどういうことか?ということを追求して品質の高いヒト組織細胞の創出に取り組んでいます.次回は,私たちの成果を含め,ヒト幹細胞を用いた応用研究のトピックスをご紹介したいと思います.
現在,私たちは,ヒトiPS細胞から創薬開発に有用な良質のヒト肝細胞を代替する細胞を沢山作り出すことを目指しています.マウスやヒトのES細胞で既に膨大な分化研究がなされているものの、その品質は生体肝細胞からは遥かに遠いものに留まっています.溢れるほどの情報を活用しながらも,組織細胞が機能的に成熟するとはどういうことか?ということを追求して品質の高いヒト組織細胞の創出に取り組んでいます.次回は,私たちの成果を含め,ヒト幹細胞を用いた応用研究のトピックスをご紹介したいと思います.
文献
- James, A. W. et al. An abundant perivascular source of stem cells for bone tissue engineering. Stem Cells Transl Med 1, 673-684, doi:10.5966/sctm.2012-0053 (2012).
- Crisan, M., Corselli, M., Chen, W. C. & Peault, B. Perivascular cells for regenerative medicine. J Cell Mol Med 16, 2851-2860, doi:10.1111/j.1582-4934.2012.01617.x (2012).
- Kuroda, Y. et al. Unique multipotent cells in adult human mesenchymal cell populations. Proc Natl Acad Sci U S A 107, 8639-8643, doi:10.1073/pnas.0911647107 (2010).
- Wakao, S. et al. Multilineage-differentiating stress-enduring (Muse) cells are a primary source of induced pluripotent stem cells in human fibroblasts. Proc Natl Acad Sci U S A 108, 9875-9880, doi:10.1073/pnas.1100816108 (2011).
- Milo, R. and Phillips, R. Cell Biology by the Numbers. 1st edition. CRC Press Book (2015).
- Takahashi, K. & Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126, 663-676 (2006).
- Evans, M. J. & Kaufman, M. H. Establishment in culture of pluripotential cells from mouse embryos. Nature 292, 154-156. (1981).
- Zhao, X. Y. et al. iPS cells produce viable mice through tetraploid complementation. Nature 461, 86-90, doi:10.1038/nature08267 (2009).
- Takahashi, K. et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 131, 861-872 (2007).
- Yu, J. et al. Induced pluripotent stem cell lines derived from human somatic cells. Science 318, 1917-1920, doi:1151526 [pii]10.1126/science.1151526 (2007).
- Hatano, S. Y. et al. Pluripotential competence of cells associated with Nanog activity. Mech Dev 122, 67-79 (2005).
- Kuroda, T. et al. Octamer and Sox elements are required for transcriptional cis regulation of Nanog gene expression. Mol Cell Biol 25, 2475-2485, doi:10.1128/MCB.25.6.2475-2485.2005 (2005).
- Niwa, H., Ogawa, K., Shimosato, D. & Adachi, K. A parallel circuit of LIF signalling pathways maintains pluripotency of mouse ES cells. Nature 460, 118-122, doi:10.1038/nature08113 (2009).
- Hishida, T. et al. Indefinite self-renewal of ESCs through Myc/Max transcriptional complex-independent mechanisms. Cell Stem Cell 9, 37-49, doi:10.1016/j.stem.2011.04.020 (2011).
- Wilmut, I., Schnieke, A. E., McWhir, J., Kind, A. J. & Campbell, K. H. Viable offspring derived from fetal and adult mammalian cells. Nature 385, 810-813, doi:10.1038/385810a0 (1997).
- Tada, M., Tada, T., Lefebvre, L., Barton, S. C. & Surani, M. A. Embryonic germ cells induce epigenetic reprogramming of somatic nucleus in hybrid cells. EMBO J 16, 6510-6520, doi:10.1093/emboj/16.21.6510 (1997).
- Tada, M., Takahama, Y., Abe, K., Nakatsuji, N. & Tada, T. Nuclear reprogramming of somatic cells by in vitro hybridization with ES cells. Curr Biol 11, 1553-1558 (2001).
- Kimura, H., Tada, M., Nakatsuji, N. & Tada, T. Histone code modifications on pluripotential nuclei of reprogrammed somatic cells. Mol Cell Biol 24, 5710-5720 (2004).
- Tada, M. et al. Pluripotency of reprogrammed somatic genomes in embryonic stem hybrid cells. Dev Dyn 227, 504-510 (2003).
- Takagi, N., Yoshida, M. A., Sugawara, O. & Sasaki, M. Reversal of X-inactivation in female mouse somatic cells hybridized with murine teratocarcinoma stem cells in vitro. Cell 34, 1053-1062 (1983).
- Thomson, J. A. et al. Embryonic stem cell lines derived from human blastocysts. Science 282, 1145-1147 (1998).
幹細胞リプログラミング研究室 多田 政子



























